Зубр. Популяционно-генетический анализ
Сипко Т. П.
Институт проблем экологии и эволюции РАН, Москва
Аннотация. Предпринят анализ истории формирования современной популяции зубра. Проведено сравнение полученных генетических и демографических характеристик между разными формами этого вида. Обсуждается изменения состояния вида во времени.
European bison. The population and genetics analysis
Sipko T. P.
Institute of Problems of Ecology and Evolution of RAS, Moscow
The analysis of the history of formation of a modern population of the European bison is undertaken. Matching the obtained genetic and demographic characteristics between the miscellaneous forms of this species is conducted. The state of transitions of the species in the course of time is discussed.
Введение. Современное состояние вида
По данным Международной родословной книги (EBPB, 2000) на 2000г. мировая популяция так называемого чистокровного зубра насчитывает 2864 особи, из них 1154 зубров содержится в 192 пунктах разведения в неволе (зоопарки, зоосады, питомники, парки) и 1710 в 33 вольных стадах и с 1987 года существенно не изменяется. А в последнее десятилетие имеется тенденция к сокращению численности этого вида.
Зубра в мире содержат в виде разобщенных относительно малочисленных популяций (в вольноживущих стадах насчитывается от 1 до 300 особей). Большинство вольных стад, а также зубры из питомников расположенных в России и странах Восточной Европы происходят только от пяти-шести особей основателей (Белоусова, 1993). И только зубры, обитающие в Кавказском Государственном заповеднике, имеют генеалогические связи как с 11 из имеющихся родоначальников кавказско-беловежской линии, так и с еще 3 особями, генеалогическая линия от которых идет только к горной линии. В процессе восстановления вид прошел, по крайней мере, через три "бутылочных горлышка" (катастрофического сокращения численности).
Более 300 лет эффективная численность зубров в Беловежской пуще, являющейся основным донором этих животных, не соответствовала параметрам позволяющим гарантировать долгосрочное сохранения генетического полиморфизма этого вида. И все родоначальники современных зубров были вывезены, для дальнейшего разведения, не из дикой популяции, а из питомника существующего при охотничьем парке «Беловежская пуща» с середины ХIХ века. В этом питомнике (Зверинце) содержали обычно 10-30 особей зубров, что неминуемо вело к росту их инбридинга.
Внушают тревогу, в целом относительно медленные темпы прироста численности в вольных стадах зубра; соответствующие показатели значительно ниже, чем, например, у канадского лесного бизона, также пережившего резкое сокращение численности, но не столь катастрофического масштаба.
Современные исследования показали значительное сокращение генетического разнообразия зубра и, как следствие, снижение его адаптационного потенциала (Сипко и др., 1993, 1994, 1995, 1996; Hartl, Pucek, 1994; Раутиан, Сипко, 1996; Baranov et al, 1997), что вызывает обоснованные опасения в отношении, возможности, выживания отдельных линий разведения зубра в будущем. Снижение индивидуального генетического разнообразия популяций и видов приводит к потере эволюционной пластичности. В результате популяции становятся неустойчивыми к действию возбудителей болезней, конкурентов, изменению окружающей среды (климат, кормовые ресурсы) и т.д. (Nevo et al., 1985; O`Brien, Evermann, 1988; Parsons, 1989; Ferguson, Drahuschak, 1990). Чтобы обеспечить краткосрочное выживание популяции, эффективная численность должна составлять примерно 50 особей (Senner, 1980; Franklin, 1980; Soule., 1980; Frankel, Soule, 1981). По этой причине питомники по разведению зубра не могут выполнять функцию по сохранению страхового генофонда зубра из-за малой численности племенного поголовья. Последнее неизбежно приводит к генетико-автоматическим процессам (эффект основателя, генетический дрейф, инбредная депрессия), естественному и бессознательному искусственному отбору, что существенно нарушает и обедняет генетический фонд этих микропопуляций (Сипко, Белоусова, 1993).
Чтобы обеспечить долгосрочное выживание популяции и непрерывное развитие адаптаций, эффективная численность должна составлять примерно 500 особей (Senner, 1980; 1980; Soule., 1980; Frankel, Soule, 1981). Это соответствует единой популяции числом в 1500-2000 особей. А реконструкция видового ареала зубра, в настоящем, и в обозримом будущем, маловероятна в связи с сильным антропогенным давлением практически на всей территории исторически сложившегося поздне-плейстоценового ареала вида. В этой связи можно рассчитывать только на создание отдельных достаточно крупных вольноживущих популяций, в которых возможно восстановление естественных генетических и биологических процессов, что было предложено как итог работы рабочей группы специалистов по зубру SSC/IUCN, специалистов по сохранению и разведению SSC/IUCN, EEP проходившей под председательством Z. Pucek, U.S. Seal и P. Miller в июне 1995.
Происхождение современных зубров
История истребления, спасения и разведения зубра хорошо документирована. Этот факт позволяет нам проследить происхождение и родословные связи зубров за большой промежуток времени. В общих чертах история зубров состоит из трех этапов. Это существование в дикой природе, и полное истребление, затем разведение в вольерах под контролем человека и реинтродукция в природу.
В голоцене и в раннеисторическое время ареал зубра занимал территорию почти всей Европы за исключением её северной части (Флёров 1979). И на этой территории морфологические, физиологические и все генетически детерминированные признаки изменялись клинально от Белорусских лесов до Кавказских предгорий. По историческим свидетельствам зубр был многочислен на северном причерноморье от Карпат и низовий Дуная до Кубани и Кавказских гор (Кириков, 1979), где вероятнее всего и имелись оптимальные для этого вида условия. И можно утверждать, что на Юге Русской равнины жил зубр, которого можно было бы назвать Беловежско-Кавказским, а с его истреблением была утрачена большая часть полиморфизма свойственная этому виду.
К началу XVIII века единый ареал зубра оказался разорванным на несколько изолированных участков (представляющих собой малопригодные для сельскохозяйственной деятельности территории). А к началу XIX века на земле вольные стада сохранились только в Беловежской пуще и на северо-западном Кавказе (Юргенсон, 1956, Pucek, 1991). Их определили как два подвида: беловежский (Bison bonasus bonasus Linnaeus, 1758), и кавказский (Bison bonasus caucasicus Satunin, 1904). Исчезновение зубра в дикой природе было вызвано причинами антропогенного характера: уничтожением местообитаний (вырубка и выжигание лесов, преобразование лесных массивов, степей и речных пойм в сельскохозяйственные угодья) и неограниченной охотой. В начале ХХ века вид был в природе уничтожен полностью, за исключением 54 экземпляров, содержавшихся в различных зоопарках Европы (Pucek, 1991). Кавказский подвид был уничтожен полностью, за исключением потомства от быка кавказского подвида (Caucasus N 100 RZR) и беловежских зубриц. Эти животные, выделяются в кавказско-беловежскую линию разведения, которая, в соответствии с Международным кодексом зоологической номенклатуры (1988) относятся к чистокровным особям данного вида.
Беловежская пуща в силу своих природных условий и исторического пути стала местом, где долгое время сохранялись вольно живущие зубры, а последний был застрелен 9 февраля 1921 г. бывшим служащим охраны Варфоломеем Шпаковичем (Башкиров, 1939; Юргенсон, 1956). И из Беловежского зверинца (вольеры) Русским Императором были дарены зубры, которые пережив разрушения Первой Мировой войны стали основой для восстановления этого вида и основателями беловежской линии разведения.
О зубре из беловежской Пущи сохранилось много публикаций (Усов, 1865; Карцев, 1903; Калугин, 1919; Wroblewski, 1927; Pucek, 1991) которые мы смогли использовать для составления этого обзора. И если с начала XIХ века мы имеем достаточно подробные данные о численности зубра в Беловежской пуще, то для предыдущих веков мы можем только использовать приблизительные цифры. В 1538 году вышел закон об охране лесов и охот в Великом Княжестве Литовском, а в 1558 году произвели описание пущи. На основании этих фактов можно уже предположить, что у современников этих законов были серьезные основания для беспокойства по состоянию охотничьей фауны и в том числе зубра. Истребление зубра привело к тому, что к 1600 г. беловежская популяция зубров стала полностью изолированной. По проведенному в 1640 году описанию пущи ее площадь составляла примерно 150 тыс. десятин, что равняется 163,5 тыс. га. И если принять, что для обитания зубра в пущи пригодно примерно 60% территории (Карцев 1903, Krasinski et. al, 1994) можно считать, что зубр занимал площадь в 98.1 тыс. га. Современные данные, по Беловежской пущи дают плотность заселения зубров в 8-11 гол на 1000 га. Таким образом, по расчетам эту территорию может населять 784-1080 голов зубров. Для примера: По описанию 1842 года территория пущи занимала 117,6 тыс. десятин, что составляет 128,2 тыс. га зубр заселял 70000 десятин (Карцев 1903) и при возможной плотности в 4-7 голов на тыс. га число зубров составляет 610-840 особей, что соответствует данным приведенным Карцевым (1903) по численности зубров в XIX веке. По данным Красинского с соавторами (1994) общая площадь, освоенная зубрами в обеих частях Беловежской пущи составила 53 тыс. га., где по расчетам может обитать 424-583 голов, что соответствует современным параметрам численности зубра в беловежской пуще. Также нужно принять во внимание, что в прошлые века значительное воздействие на зубра оказывали законные и не законные охоты, и заметно влиял пресс хищников (Карцев 1903), а, также учитывая тот факт что, Беловежская пуща никогда не являлась оптимальной территорией для обитания зубра (Кулагин, 1919). В то время пуща была более заболочена и покрыты старым лесом, т.е. представляла собой угодья не позволяющие обеспечить кормом значительное число постоянно обитающих на этой территории крупных животных. Нельзя забывать, что на период между XVII-XIX веками приходится вторая половина малого ледникового периода (Кренке, 1995) характеризуемая низкими температурами высоким и долго лежащим снеговым покровом. Таким образом, по нашим оценкам, территорию Беловежской пущи в 1600-1800 годы могло населять не более 800 особей.
Пик численности зубров приходящийся на 1830-1860 годы в беловежской пуще и описываемый Карцевым (1903) и Кулагиным (1919) основывается на информации, полученной из ведомостей, которые велись в то время в пуще. Усов (1865), Карцев (1903), Северцев (1940) анализируя, имеющиеся данные по динамике зубров за этот период высказывает обоснованные сомнения в достоверности этой информации. Приводимые расчеты также не позволяют надеяться, что такая территория могла обеспечивать жизнь 1898 особей зубра. Также необходимо принять во внимание, восстания сепаратистов в 1830-31 и 1863 годах, в результате которых отсутствовала надлежащая охрана зубров, и процветало браконьерство. Таким образом, мы разделяем точку зрения Карцева, что в период с 1830 по 1864 годы реальное число зубров не превышало данных по 1829 и 1865 годов, что составило 711 и 724 особей соответственно. А это значит что более 300 лет (примерно 60 поколений) эффективная численность популяции зубра в Беловежской пуще была значительно меньше предложенной нижней границы численности необходимой для сохранения разнообразия вида в долговременном плане и возможности эволюции в меняющейся среде (Франклин, 1980).
Истребление зубров обитающих на Кавказе проходило несколько медленнее, чем в Европейской части ареала. Это было вызвано в первую очередь многовековым противостоянием между кочевниками и горцами. В результате чего горцы вынуждены были, селится в труднодоступных ущельях. А богатый по своим природным условиям район между предгорьями северного макросклона Кавказского хребта и поймами таких рек как Кубань и Терек был практически лишен постоянного оседлого населения. Этот факт позволил пережить кавказским зубрам своих родственников из многих других районов.
Ситуация с сохранением зубра стала меняться в худшую сторону с началом освоения Предкавказья Россией. Когда в результате наведения относительного порядка в этом регионе, резко сократилось число междоусобных войн, и прекратилась работорговля. И как следствие этот регион, получив такую возможность, стал интенсивно обживаться и осваиваться. Возник постоянный пресс на популяцию, которая стала заметно сокращаться. Несколько замедлилось истребление зубра в период «Кавказских войн» происходивших в середине XIX века и отвлекавших взрослое мужское население от эксплуатации природных ресурсов.
Начавшаяся после окончания «Кавказской» войны в 60-х годах массовая миграция населения в этом регионе в первое время отвлекала людей от истребления животных. А по мере цивилизации и освоении региона ареал зубра продолжил сокращаться. И запрещение охоты на зубра с 1867 года и так восторженно принятая натуралистами, организация в 1888 году в районе обитания кавказского зубра, охотничьего хозяйства «Кубанская охота» площадью в 522 тыс. га. принадлежащего царской фамилии явились, к сожалению, лишь отсрочкой его гибели. В 1917 году «Кубанская охота» была национализирована, а затем Гражданская война, браконьерство, эпизоотии, беззаконие и зубр на Кавказе исчез.
В шестидесятые годы XIX века, сохранившиеся на северо-западном Кавказе зубры, получили возможность наблюдаться учеными. И имеются публикации с оценкой числа зубров на Кавказе в разные годы. Мы себе отдаем отчет в том, что адекватной методики учета численности зубров в труднопроходимой, горно-лесной местности тогда не существовало и в том, какие трудности стояли перед первыми исследователями. Поэтому приведенные в таблице 1 данные предлагаем использовать как оценочные.
Таблица 1. Динамика численности зубра на Кавказе до его истребления
| Годы | Число | Источник информации |
| 1868 | 2000 < | Westberg, 1899 |
| 1871 | 2000 | Westberg, 1895 |
| 1880 | 500< | Башкиров, 1939 |
| 1890 | 1000 | Westberg, 1895 |
| 1893 | 800 | Westberg, 1899 |
| 1894 | 700 | Шильдер, 1895, Сатунин,1898 |
| 1910 | До 600 | Калугин, 1919; Pfizenmayer, 1926 цит. По Башкирову, 1939 |
| 1917 | До 500 | Силантьев, 1919 |
| 1919 | До 100 | Шапошников, 1928 |
| 1920 | 20< | Розанов, 1928 |
| 1925 | 15-20 | Фортунатов, 1932 |
| 1927 | --- | Пастухами имеретинами убиты последние 3 зубра Башкиров (1939) по данным наблюдателя охраны А.П. Онешко |
Сохранились известия о трех попытках вывести зубра с Кавказа. Бык 1866 года рождения пойманный в урочище Верхний Уруп в 1867 году был привезен в Московский зверинец. Бык, родившийся в мае 1899 года в урочище Псебай в октябре того же года, был доставлен в Беловежский зверинец, где, как точно известно, жил несколько лет. А для нас наибольший интерес представляет третье животное, которому суждено сыграть заметную роль в спасении зубров от вымирания как вида и единственного оставившего гены кавказского зубра. Эта особь была изъята из популяции еще не испытавшей значительного сокращения численности (см. таблицу 1) т.н. стадии «бутылочного горлышка» поэтому с большой долей вероятности можно считать что это аутбредное животное с коэффициентом инбридинга около нуля что и подтверждается нашими расчетами (Табл. 2). Пойманный, в мае 1907 года на склоне горы Пшекиш, в трехмесячном возрасте зубренок был перевезен в Беловежскую пущу, а в 1908 году Император России подарил его Гагенбеку у которого в городе Гамбурге он жил до 1922 года. Затем Гагенбек уступил зубра графу Арниму, и его перевезли в Бойтценбург (Boitzenburg) где он дожил до естественной смерти 26 февраля 1925 года. Этот бык получил кличку Кавказ и номер 100 в племенной книге «EBPB» и за свою жизнь оставил только 7 телят (3 быка и 4 коровы), от беловежских зубриц которые и стали основателями линии разведения зубров получившей название «кавказско-беловежская линия».
Одной из особенностей происхождения практически всех основателей современных линий разведения, не замеченной многими авторами, является тот факт, что эти зубры и их предки не были отловлены из дикой природы, а были вывезены из питомника существующего, но территории Беловежской пущи с 1860 года (Карцев, 1903). Там десятилетия они разводились «в себе» и при небольшой численности в пределах 6-32 особей. Таким образом, уже в питомнике проходило близкородственное разведение и как результат рост инбридинга, отрицательные последствия которого уже тогда вызывали необходимость в «освежении крови» для чего из Гатчинского зверинца привозили зубров и до нас дошли сведения о завозе в 1900 году 4 зубров (Юргенсон и др. 1956).
Для получения данных по коэффициенту инбридинга основателей мы предпринял моделирование родословных этих животных исходя из имеющихся литературных данных по Карцеву (1903) и минимизируя число поколений в спорных случаях. Средний коэффициент инбридинга зубров Беловежской пущи в 1860 году, на момент основания Беловежского питомника по расчету, проведенному из эффективной численности (Сулей, 1983) составил 0.093. По той же формуле рассчитывались показатели инбридинга и в питомнике. Полученные значения представлены в таблице 2.
Таблица 2. Значение коэффициента инбридинга зубров родоначальников современных линий разведения.
| Номер по племенной Книге EBPB | Кличка | коэффициент инбридинга |
| 15 | Бегрюндер | 0.223 |
| 16 | Плавия | 0.198 |
| 35 | Плевна | 0.198 |
| 42 | Планта | 0.198 |
| 45 | Плебейер | 0.237 |
| 46 | Плацида | 0.237 |
| 50 | Белосток | 0.256 |
| 51 | Биала | 0.256 |
| 87 | Биль | 0.366 |
| 89 | Бильма | 0.311 |
| 95 | Гарде | 0.285 |
| 96 | Гатчина | 0.311 |
| 100 | Кавказ | 0.015 |
| 147 | Бисмарк | 0.403 |
Выведение «Горной» линии зубров было вызвано желанием воссоздать или создать популяцию зубра на Кавказе в замен истребленного подвида. Задача осложнялась тем, что когда возникла в этом необходимость, в СССР был всего один чистокровный зубр самец и небольшое число помесей между зубрами и бизонами. Приобретение дополнительно зубров в то время было не реально по политическим и экономическим причинам.
Селекционная работа по выведению зубра пригодного к обитанию в сложных условиях сильно пересеченной горно-лесной местности проводилась по этапам.
I. Гибридизация между зубрами и бизонами с 1905 по 1935 годы
II. Поглотительное скрещивание имеющейся группы помесей на зубра с использованием быка Бодо 193 с 1935 по 1940 годы.
III. Перемещение группы в 5 особей – интродуцентов в район натурализации и разведение «в себе» 1940-1949 годы.
IV. Повышение кровности стада по зубру. Проводилось с привлечением для воспроизводства чистокровных зубров кавказско-беловежской линии (реально участвовало в размножении 15 особей) при полном отстранение от размножения гибридных самцов 1949-1959 годы.
V. С 1960 года племенной учет прекращен в связи с полном переходом зубров к дикому образу жизни. В размножении стали эффективно участвовать самцы гибридогенного происхождения. Доля крови бизонов становиться равной в среднем 5.6%* (Сипко, 1990). Племенная работа стала заключаться в селекционном отстреле.
Селекция в Кавказском заповеднике проходила параллельно с работами по адаптации и натурализации, животных на Кавказе. По данным Калугина (1958, 1965) в 1940-45 годы практиковалась загонная система содержания животных. В период 1946-1953 гг. зубры содержались по подкормочно-выпасной системе, когда в течение 240-300 дней они свободно паслись, получая лишь минимальную подкормку в виде поваренной соли, а на 2-3 месяца переводились на искусственную подкормку. С 1949 по 1954 годы включительно производили перегон зверей на высокогорные пастбища, а затем они стали проделывать эти миграции самостоятельно. И с 1954 года необходимость подкормки животных отпала в связи с их полным одичанием. Сроком начала вольного разведения зубра на Кавказе правильно считать 1949 год, когда эти звери уже большую часть года стали обходиться без помощи человека.
Распределение генетического пула родоначальников в современных зубрах
Исторически сложилось разведение зубра по линиям. Это беловежская линия, происходящая от 5 основателей, кавказско-беловежская, которая происходит от 12 животных-родоначальников, включая и основателей беловежской линии, уцелевших после истребления вида к началу ХХ века. И горная линия, получившая статус подвида (Раутиан и др.,2000). В каждой из современных популяций представлена только часть генетического материала, существовавшего во времена расцвета вида (Сипко и др, 1993, 1994, 1995, 1996).
Как видно из таблицы 3, вклад родоначальников в генофонд зубров крайне неравномерный и хотя для питомников России он в основном и соответствует пропорциям в средне мировой популяции, можно заметить низкую долю генотипов от некоторых родоначальников. Но надо отметить, что пока в части пунктов разведения (Табл. 5), пропорции доли геномов предков несколько иные и это еще является ресурсом для работы по сохранению генетического разнообразия этого вида.
Таблица 3. Генетический пул основателей (в %) зубров в питомниках России*
| Основатели | 42,45 | 15,16 | 147 | 95 | 96 | 100 | 35 | 46 | 87,89 |
| ОГЗ, F КБЛ, M Cp. |
53.5 52.2 53.1 |
11.3 10.9 11.1 |
0 0 0 |
2.7 2.6 2.6 |
5.3 5.3 5.3 |
2.8 2.8 2.8 |
0.1 0.2 0.1 |
0 0 0 |
18.9 18.9 18.9 |
| ПТЗ, F КБЛ, M Cp. |
53.8 51.6 53.2 |
12.3 13.0 12.5 |
0.1 0 0.1 |
2.7 3.0 2.8 |
5.3 5.4 5.3 |
3.2 5.5 3.8 |
0.1 2.5 1.1 |
0.07 0.58 0.21 |
18.8 18.6 18.7 |
| ПТЗ, F БЛ, M Cp. |
94.4 95.3 94.6 |
1.3 1.3 1.3 |
0.4 0.4 0.4 |
0 0 0 |
0 0 0 |
0 0 0 |
0 0 0 |
0 0 0 |
3.8 3.0 3.6 |
| В мире** | 46.3 | 14.8 | 0.6 | 3.5 | 5.7 | 6.7 | 3.2 | 1.3 | 17.8 |
* В питомнике "Черга" вклад основателей практически идентичен с беловежской
линией из Приокско-террасного заповедника.
** в среднем по всем питомникам в мире (Olech, 1989)
Также можно отметить, что в процессе разведения зубров из имеющихся 5 быков основателей уже прервалась отцовская линия, а значит наследование половой «Y» хромосомы у основателей N 87; 147 и 15. Это грозит и для основателя N 100 быка по кличке Кавказ уже в ближайшее время, что значительно обеднит и так усеченный генофонд этого вида.
Как уже сообщалось, питомники по разведению зубра не могут выполнять функцию сохранения страхового генофонда зубра из-за малой численности племенного поголовья. Так как длительное содержание в питомнике большого поголовья животных не позволительно по экономическим причинам. Но в тоже время питомники могут и должны, выполнять следующие функции: а) адаптация зубров выросших в зоопарках к условиям вольного обитания; б) реинтродукция криоконсервированных геномов в популяцию; г) размножение редких по генеалогическим и генетическим показаниям групп зубров. Создание генетического криобанка позволяет при его маленьком объеме иметь большую и репрезентативную коллекцию криоконсервированных геномов. Их можно хранить неограниченно долго и легко перевозить на любые расстояния. К нашему сожалению это направление не получает должной поддержки. Хотя методика получения и криокорсервирования половых клеток зубра разработана (Сипко и др., 1993), апробирована (Sipko et al., 1997) и служит для проведения работ по межвидовой гибридизации (Абилов и др., 1994). В настоящее время в генетическом криобанке хранятся криоконсервированные мужские половые клетки 11 особей зубра.
Семя от зубра можно получать и тогда, когда он подлежит выбраковке по селекционным или ветеринарным показаниям. Использование криоконсервированного семени через два и более поколения позволяет увеличивать и поддерживать комбинативную изменчивость. При этом достаточно просто осуществляется её транспортировка.
Для реинтродукции геномов сохраняемых в криобанке необходимы питомники для разведения зубров, где содержаться животные реципиенты которых можно использовать для искусственного осеменения. Полученные в результате этих операций особи, несущие половину генов от донора, можно направлять как на разведение, так и для расселения в вольных популяциях.
Отмечалось выше, в современных популяциях крайне неравномерно представлен генетический пул основателей. Особенно это характерно в вольных стадах. Мы рассчитали их вклад для животных, выпущенных в природу (Табл. 4, 5). Можно видеть в таблице 5, что основатели так называемой Плесской линии (NN 42, 45) внесли наибольший вклад в генотипы выпущенных животных (больше половины), а для некоторых группах он больше 90%. Вместе с тем, невелик вклад прочих основателей. Можно также видеть, что имеющийся генофонд в питомниках представлен хаотично и не полно. Нет тенденции к выравниванию генетического пула основателей, а также к репрезентативному отображению имеющегося генофонда зубров. Необходимо отметить малое число основателей и популяций поставщиков животных, что не позволяет надеяться на полноценное сохранение генетического разнообразия. Для пополнения вновь создаваемых вольных популяций зубров нужно использовать животных из вольноживущих популяций (Табл. 4, 5), имеющих относительно высокую численность и приемлемый генетический полиморфизм. Организуя разведение редких племенных и генетических линий зубров в питомниках целесообразно затем использовать их при создании вольноживущих стад.
Таблица 4. Количество и источник происхождение животных, давших начало крупнейшим вольным популяциям зубров в мире
| Популяция | Завезено зубров | Годы завоза зубров | Питомник поставщик животных | |
| всего | самок | |||
| Кавказский Заповедник | 20 | 4 | 1940-1959 | Аскания-Нова, ПБП, ПТЗ |
| Нальчикское охотхозяйство, Кабардино-Балкария | 35 | 17 | 1959-1967 | КГЗ, ПТЗ, ОГЗ, БП |
| Цейский заказник и Северо-Осетинсий заповедник | 48 | 32 | 1964-1968 | ПТЗ, БП |
| Тебердинский заповедник, Архызский участок | 29 | 13 | 1968-1981 | ПТЗ, ОГЗ |
| Беловежская пуща, Беларусь | 49 | 18 | 1964-1991 | БП, ПТЗ, ПБП |
| Беловежская пуща, Польша | 38 | 24 | 1952-1966 | ПБП |
| Буковинское и Зубровица о/х | 23 | 14 | 1970-1977 | БП, ПТЗ, ОГЗ |
| Бещады | 63 | 29 | 1963-1982 | ПБП |
| Цуманское и Клеванское о/х | 23 | 14 | 1965-1968 | БП |
Примечание: БП - Беловежская пуща (Беларусь); ПБП - Беловежская пуща (Польша); КГЗ - Кавказский государственый заповедник; ОГЗ - Окский государственый заповедник; ПТЗ - Приокско-Террасный государственый заповедник.
Таблица 5. Генетический пул (“доля крови”) основателей в современных популяциях зубра (вольных популяциях, а также основных зарубежных центрах разведения зубров), %
| Популяция | Номера зубров-основателей по EBPB | Бизон B.bison | ||||||||
| 45, 42 | 89, 87 | 15, 16 | 123, 122 (126, 147) | 100 *** | 96, 95 | 32 , 33 (35, 36) | 46 | 50, 51,65 | ||
| Кавказский заповедник | 31.40 | 17.63 | 18.58 | 1.86 | 5.29 | 10.42 | 1.81 | 0 | 6.61** | 6.40 |
| Цейский заказник | 62.79 | 18.34 | 9.42 | 0.19 | 2.25 | 6.76 | 0 | 0 | 0 | 0 |
| Тебердинский заповедник | 61.25 | 18.91 | 9.92 | 0 | 2.48 | 7.44 | 0 | 0 | 0 | 0 |
| Буковинское хозяйство | 62.80 | 18.94 | 9.59 | 1.06 | 2.32 | 6.96 | 0 | 0 | 0 | 0 |
| Цуманское хозяйство | 58.81 | 20.54 | 0.70 | 0 | 2.67 | 8.02 | 0 | 0 | 0 | 0 |
| Великоозерское хозяйство | 94.29 | 2.98 | 1.90 | 0.88 | 0 | 0 | 0 | 0 | 0 | 0 |
| Усть-кубенское хозяйство | 75.43 | 11.96 | 7.10 | 0.20 | 1.32 | 3.94 | 0 | 0 | 0 | 0 |
| Фоминское | 55.27 | 18.80 | 13.35 | 0 | 3.45 | 8.35 | 0.62 | 0.15 | 0 | 0 |
| Брянский лес | 57.56 | 20.07 | 10.73 | 0 | 3.08 | 8.19 | 0.29 | 0 | 0 | 0 |
| Орловское полесье на 1999 г. | 76.32 | 6.77 | 9.09 | 0.06 | 1.97 | 5.80 | 0.05 | 0 | 0 | 0 |
| Скнятинское хозяйство | 57.15 | 20.03 | 11.05 | 0.02 | 2.64 | 7.73 | 0.06 | 0 | 0 | 0 |
| Польша, Беловежская пуща | 73.30 | 8.40 | 13.80 | 4.60 | 0 | 0 | 0 | 0 | 0 | 0 |
| Беларусь, Беловежская пуща | 78.77 | 11.82 | 6.37 | 2.65 | 0 | 0 | 0 | 0 | 0 | 0 |
| Авеста (Avesta)* | 25.9 | 33.4 | 16.7 | 0 | 6.9 | 15.2 | 1.8 | 0 | 0 | 0 |
| Шпринге (Springe)* | 35.1 | 22.3 | 18.4 | 2.4 | 7.0 | 9.9 | 3.0 | 2.1 | 0 | 0 |
| Harde-hausen* | 32.9 | 8.4 | 18.4 | 1.1 | 13.8 | 11.7 | 8.0 | 5.8 | 0 | 0 |
| Damerover Werder* | 49.5 | 27.6 | 10.8 | 0 | 3.2 | 8.6 | 0.3 | 0 | 0 | 0 |
| Tpocianky* | 26.6 | 28.9 | 9.8 | 0 | 4.3 | 9.2 | 1.2 | 0 | 0 | 0 |
| Whipsnade* | 56.3 | 13.1 | 14.7 | 2.5 | 4.8 | 5.3 | 2.8 | 0.7 | 0 | 0 |
| Sabadurg* | 47.3 | 21.1 | 12.6 | 0 | 6.0 | 9.8 | 2.4 | 0.9 | 0 | 0 |
| В мире* | 46.3 | 17.8 | 14.8 | 0.6 | 6.7 | 9.2 | 3.2 | 1.3 | 0 | 0 |
*по W. Olech (1989) остальные оценки получены нами на основе родословных книг (EBPB).
** Из них 5.18% унаследовано от Белостока (EBPB N 50), 0.73% ? от Биалы (EBPB N 51) и 0.71% от Петера (EBPB N 65).
*** По данным Государственной племенной книги (ГПК, 1956), во второй половине XIX века было пойман, доставлен и Беловежскую пущу живой зубренок кавказского подвида. Этот бычок, родившийся 12 мая 1899 года, 25 мая был доставлен в вольеру п. Псебай, а в октябре того же года в Беловежский зверинец. В зверинец этот бык был привезен специально для «освежения крови» и известно по Карцеву (1903), что он жил там более 4 лет под именем Казан (ГПК N 58). Сведений об участии быка Казана в скрещиваниях не сохранилось. Однако, в то время подробных племенных записей не вели, а за время Первой мировой войны все документы были утеряны. Принято считать, что Казан не оставил потомства (Башкиров, 1939); во всяком случае, его генетический вклад в популяцию современных зубров никогда не брался в расчет. Но с другой стороны трудно исключить факт использования этой особи в разведении, а тем более в условиях питомника, куда он и был помещен с этой целью. Принимая эти рассуждения можно предполагать, что зубры, родившиеся в беловежском зверинце, после 1903 года могут являться детьми быка Казана и иметь «кровь» кавказского подвида. Наиболее вероятно это зубры, ставшие, в дальнейшем, родоначальниками современных линий Бибер № 123, Биберштеин № 85, Бильдунг № 86, Бильма № 89, Гарде №95, Гатчина № 96.
Из питомника Беловежской пущи на волю в 1963 году выпустили 3 особи с неполными племенными записями. В то время в Беловежском питомнике имелись 35 животных также с не полностью известными родословными, которых относили к кавказско-беловежской линии, то вполне возможно, что потомки основателей кавказско-беловежской линии могут быть представлены в популяции зубров Беловежской пущи.
Зубр бык №1229 Pomyslny (родители - f902 Ponury и m736 Podwika) относимый к беловежской линии родился в 1959 году в Польше и в 1963 г. передан в Беловежскую пущу. Но в племенной книге за 1965-69 гг., на 7 странице (глава «Additions and corrections to former liste») записана матерью этого животного корова № 979 Purella. Этот бык был выпущен на волю в Белорусскую часть Беловежской пущи в мае 1963 года. В случае если это не очередная ошибка, в вольную популяцию Беловежской пущи выпустили зубра, относящегося к кавказско-беловежской линии.
Анализ истории формирования популяции зубра в беловежской пуще показал, что на территории пущи остались в 60-х годах ХХ века 4 особи зубра относящегося к кавказско-беловежской линии (Буневич, 2003). Принимая во внимание тот факт, что только в 1981 году была установлена система пограничных инженерных сооружений, разделившая беловежский лес и популяцию зубра на две части можно экстраполировать сомнение в генетической «чистоте» зубров на всех животных обитающих в данном регионе.
Витальные характеристики зубров
Современные чистокровные зубры, относящиеся, к беловежской и кавказско-беловежской линиям разведения отличаются друг от друга по ряду важных характеристик. Из таблицы 6, можно видеть, что беловежская линия уступает по всем параметрам жизнеспособности и плодовитости. Изучение динамики витальных характеристик (на примере зубропитомников Окского и Приокско-Террасного заповедников) показывает общий тренд на ухудшение всех этих параметров (Табл. 7, 7а). В целом для мировой популяции витальные характеристики также имеют тенденцию к ухудшению, но не достоверно (Табл. 7б).
Динамика витальных характеристик в горной линии, не относящейся к чистокровной, (Табл. 7в) определяется кардинальными изменениями условий своего существования и отличается от других линий зубров. В 1901-1940 годы проводилась гибридизация зубров с бизонами и разведение в степных условиях Аскании-Нова. С 1941 по 1950 годы эти животные были доставлены в вольеры расположенные на горе, где начался процесс интродукция и адаптации к условиям Кавказского заповедника при практике - разведения “в себе”. В 1951-1960 годы адаптация животных к условиям гор продолжалась, и уже большую часть времени животные добывали корм самостоятельно, отмечен в это время рост численности, а также осуществлялся завоз 15 особей новых производителей. С 1961 года полный переход на вольное содержание животных и прекращение ведения племенных записей.
Сложившаяся картина среди чистокровных зубров связана с неуклонным нарастанием во времени уровня инбридинга и сопровождается падением разнообразия. Одним из явных факторов ускоряющих этот негативный процесс является крайне неравномерное участие самцов в репродукции. Анализ вклада самцов и самок в воспроизводство (на примере всей истории существования Окской популяции) показывает, что более половины потомков получено всего от трех самцов (Табл. 8). Вклад самок в воспроизводство (Табл. 9) более равномерный, что лучше способствует сохранению имеющегося разнообразия.
Для сравнения распределения уровня смертности по возрастным классам мы привлекли данные по овцебыку. Становление популяции овцебыка на острове Врангеля проходило как при отсутствии хищников, так и преследования со стороны человека, в условиях ограниченной территории острова, то есть в условиях имеющих значительное сходство с жизнью зубров по этим параметрам, но конечно в более суровом климате. У овцебыка во многом схоже с зубром и репродуктивное поведение, социальная организация и общая продолжительность жизни. По этой причине мы предлагаем сравнить параметры их смертности в возрастных группах (таблица 10). По популяции овцебыков о-ва Врангеля данные приводятся за всю историю её существования, а для характеристики смертности зубров, данные обо всех зубрах живших в первую половину ХХ века, когда сохранению этого вида оказывалось наибольшее внимание. Хорошо прослеживается, что смертность у зубров выше во всех возрастных классах, а это может быть только результатом инбредной депрессией (Сипко и др.1999).
Таблица 6. Витальные характеристики беловежской (БЛ) и кавказско-беловежской (КБЛ) линий
| Линия | Kоэффициент инбридинга | Параметры жизнеспособности и плодовитости | ||||
| В мире | В России | Перинатальная смертность, % | Плодовитость самок, % | Возраст наступления половой зрелости, лет | интервал между отелами, месяцы | |
| Оlech, 1989 | Белоусова, 1993 | Корочкина, 1971 | Сипко, Раутиан, 1997 | |||
| БЛ КБЛ |
0,324 0,193 |
0,375 0,269 |
24,9 11,6 |
47,9 73,8 |
4,5 3,9 |
17,8 16,5 |
Таблица 7. Динамика жизнеспособности и плодовитости зубров в Окском питомнике
| Годы рождения | Родилось телят | Погибло телят | Интервал между отелами (месяцев) | Возраст первого покрытия (мес.) | Возраст первого отела (мес.) | ||||
| n | % | Сред. | Lim. | Сред. | Lim. | Сред. | Lim. | ||
| 1960-65 1966-70 1971-75 1976-80 1981-85 1986-90 1991-95 |
28 60 60 55 36 36 23 |
3 7 16 15 10 16 10 |
10,7 11,7 26,7 27,3 27,8 44,4 43,5 |
14 13 15 16 19 21 21 |
11-21 20-24 10-30 10-34 12-22 10-36 12-26 |
37,7 34,4 42,6 52,0 |
26-62 25-56 36-61 41-52; |
45,7 46,4 50,7 60,5 |
34-71 34-65 48-70 60-61 |
| Всего\Сред. | 298 | 77 | 25,8 | 16,3 | 10-36 | 39,6 | 25-62 | 48,1 | 34-71 |
Таблица 7а. Динамика изменения смертности и интервалов между отелами среди зубров Окского (ОГЗ) и Приокско-Террасного (ПТЗ) питомников.
| Годы рождения | Дожившие до репродуктивного возраста | Интервал между отелами (месяцев) | ||||||||||
| БЛ ПТЗ | КБЛ ПТЗ | КБЛ ОГЗ | БЛ ПТЗ | КБЛ ПТЗ | КБЛ ОГЗ | |||||||
| N | % | N | % | N | % | N | % | N | % | N | % | |
| 1950-65 1966-75 1976-85 1986-96 |
56 51 55 45 |
60,7 72,5 63,6 53,3 |
75 58 68 47 |
80,0 82,8 66,2 48,9 |
28 120 93 60 |
89,3 80,8 71,0 55,0 |
85 40 47 |
16,4 18,0 20,0 |
114 51 31 |
14,5 16,0 18,8 |
141 55 58 |
14,5 17,1 20,7 |
Таблица 7б. Динамика репродуктивных параметров в ХХ веке*
| 1901-1950 годы | N | 1951-2000 годы | N | |
| Зубры Беловежской линии | ||||
| Число отелов на одну самку | 5.3 | 82 | 4.9 | 89 |
| Возраст первого отела (мес.) | 56.4 | 50.1 | ||
| Юнивинальная смертность, % | 17.2 | 248 | 15.7 | 553 |
| Постнатальная смертность, % | 33.7 | 28.0 | ||
| Зубры Кавказско-беловежской линии | ||||
| Число отелов на одну самку | 5.3 | 72 | 5.5 | 264 |
| Возраст первого отела (мес.) | 47.8 | 41.0 | ||
| Юнивинальная смертность, % | 16.9 | 218 | 13.7 | 1661 |
| Постнатальная смертность, % | 27.9 | 38.0 | Бизоны в Аскании-Нова | |
| Число отелов на одну самку | 4.5 | 17 | 4.2 | 53 |
| Возраст первого отела (мес.) | 63.8 | 49.5 | ||
| Юнивинальная смертность, % | 28.1 | 32 | 14.5 | 256 |
| Постнатальная смертность, % | 37.5 | 25.0 | ||
* В расчетах до 1950 года использовались все данные по зубрам, жившим в то время, а в последующее время только по основным питомникам.
Таблица 7в. Динамика репродуктивных параметров при становлении горной линии зубров
| Годы | N | Смертность | N | Среднее число отелов на самку, особей | Возраст 1 отела, месяцев | |
| Юн % | Всего % | |||||
| 01-40 | 77 | 20.8 | 31.2 | 22 | 5.9 | 52.7 |
| 41-50 | 49 | 28.6 | 38.3 | 16 | 7.8 | 36.7 |
| 51-60 | 251 | 3.6 | 12.4 | 39 | 2.3 | 43.2 |
Таблица 8. Вклад самцов зубров из ОГЗ в воспроизводство (n=18)
| Кличка, номер | n | % | Кличка, номер | n | % |
| Муромец 1043 | 56 | 18,8 | Мещерец 1499 | 11 | 3,7 |
| Белкин 2021 | 55 | 18,5 | Мушан 2906 | 3 | 1,0 |
| Метеор 1624 | 46 | 15,4 | Мер 6183 | 3 | 1,0 |
| Мерцатель 2709 | 25 | 8,4 | Металур 5912 | 2 | 0,7 |
| Менуэт 4052 | 25 | 8,4 | Мотылек 109 | 1 | 0,3 |
| Мурманск 1241 | 24 | 8,2 | Мельник 2161 | 1 | 0,3 |
| Мушат 3831 | 20 | 6,7 | Мучек 407 | 1 | 0,3 |
| Муштабель 5642 | 14 | 4,7 | Мед 6185 | 1 | 0,3 |
| Месяму 5417 | 13 | 4,4 | Мерольд 6465 | 1 | 0,3 |
| Итого | 298 | 100 |
Таблица 9. Вклад самок зубров из ОГЗ в воспроизводство (n=43)
| Число самок | число отелов | Число самок | число отелов |
| 1 | 17 | 2 | 7 |
| 3 | 14 | 1 | 6 |
| 1 | 13 | 2 | 5 |
| 3 | 12 | 5 | 4 |
| 1 | 11 | 7 | 3 |
| 4 | 10 | 1 | 2 |
| 4 | 9 | 5 | 1 |
| 3 | 8 |
Таблица 10. Распределение смертности по возрастным группам в популяции овцебыка о. Врангеля (за 25 лет) и зубра (за 31 год)
| Возраст | Овцебык | Зубр | ||
| N | % смертности | N | % смертности | |
| 0-2 месяца | - | 4.35 | 466 | 17.1 |
| От 2 месяцев до года | 87 | 5.40 | 338 | 15.7 |
| 1 год | 344 | 3.78 | 330 | 15.8 |
| 2 года | 302 | 1.32 | 278 | 9.35 |
| 3 года | 248 | 2.42 | 252 | 9.52 |
| 4-5 лет | 351 | 2.85 | 434 | 9.22 |
| 6-7 лет | 217 | 3.23 | 349 | 9.46 |
| 8-9 лет | 143 | 4.20 | 275 | 13.1 |
| 10-12 лет | 105 | 8.57 | 292 | 11.9 |
| 13-15 лет | 35 | 31.43 | 261 | 11.5 |
| 16 и больше лет | 3 | 100 | 231 | 100 |
Генетический полиморфизм
Результаты наших исследований по генетическому полиморфизму зубра отражены в ряде публикаций (Сипко и др., 1989, 1993, 1994, 1995, 1996, 1997; Удина и др., 1994), поэтому мы не приводим характеристики использованного материала и методов исследования и рассмотрим только вопросы об уровне внутрипопуляционного разнообразия и межпопуляционной дифференциации зубра. В таблице 11 представлены характеристики иммунологического, биохимического и молекулярного полиморфизма в беловежской, кавказско-беловежской и горной линиях разведения. Все они согласно указывают на более низкий уровень разнообразия беловежской линии, который и можно было ожидать на основе данных о происхождении и современных витальных (Табл 6, 7) характеристиках двух линий разведения. Оценки полиморфизма групп крови в различных субпопуляциях с помощью mu-критерия, предложенного Л.А .Животовским (1991) приведены в таблице 12 и по этой таблице видно, что кавказско-беловежские зубры из Приокско-Террасного заповедника и Ивано-Франковска, также как и суммарная их выборка по этой линии, характеризуются более высокими значениями mu-критерия, в сравнении с беловежскими зубрами. Эти результаты отражают большие потери внутрипопуляционного полиморфизма у беловежских зубров в связи с более высоким инбридингом; вероятно данные для зубров Окского заповедника указывают на возможное обеднение их разнообразия в результате генетического дрейфа. Сопоставимые показатели для отдельных стад крупного рогатого скота (рассчитанные нами по литературным данным о 41 стаде) значительно выше, чем для зубров и составляют 1.65-1.85.
Таблица 11. Характеристика разнообразия среди современных линий зубра.
| Параметр | Линии разведения | ||
| Беловежская | Кавказско-беловежская | Горная | |
| Данные по группам крови: Количество антигенов, в линии Количество антигенов, (на особь) Разнообразие (mu-критерий) |
33 (66%) 11.2 (5-17) 1.31 |
41 (82%) 13.4 (5-18) 1.45 |
35 (70%) 12,9 (6-26) 1.50 |
| Биохимический полиморфизм: Средняя гетерозиготность (21 локус) |
0.038 | 0.044 | 0,038 |
| ДНК полиморфизм: Гетерозиготность Доля полиморфных локусов |
0,07 0,42 |
0,09 0,57 |
- - |
Таблица 12. Значения mu-критерия в разных популяциях зубра (по 54-м полиморфным генетическим маркерам)
| Популяции | Значения mu-критерия |
| Кавказско-беловежские: | |
| ПТЗ | 1.50 |
| ОГЗ | 1.40 |
| Ивано-Франковск | 1.49 |
| Суммарно | 1.50 |
| Беловежские: | |
| ПТЗ | 1.43 |
| Беловежская пуща | 1.35 |
| Суммарно | 1.39 |
| Горные: | Нальчикское ГЛОХ | 1,29 | Кавказский заповедник | 1,54 |
| Суммарно | 1,53 |
| Крупный рогатый скот (размах вариации по 41 стаду) | 1.65-1.85 |
Для характеристики генетической дифференциации, данные о генетических расстояниях между отдельными субпопуляциями, были обработаны методом многомерного шкалирования (Рис. 1). В целом генетическая дифференциация невелика, и расстояния между субпопуляциями, относящимися к одной линии разведения, сравнимы с расстояниями между субпопуляциями разных линий. Наиболее близки друг к другу кавказско-беловежские и беловежские зубры из ПТЗ, что очевидно связано с периодическим скрещиванием между ними, практикующемся в практике разведения кавказско-беловежской линии. Интересно, что горные зубры (несущие 5% примеси "крови" бизона и приведенные здесь для сравнения) несколько обособлены, но также близки к остальным популяциям (левая часть Рис. 1). Таким образом, все точки выглядят как субпопуляции единой линии; величины генетических расстояний сравнимы с таковыми для отдельных стад одной породы крупного рогатого скота (а не разных пород скота) (Сипко и др., 1995).
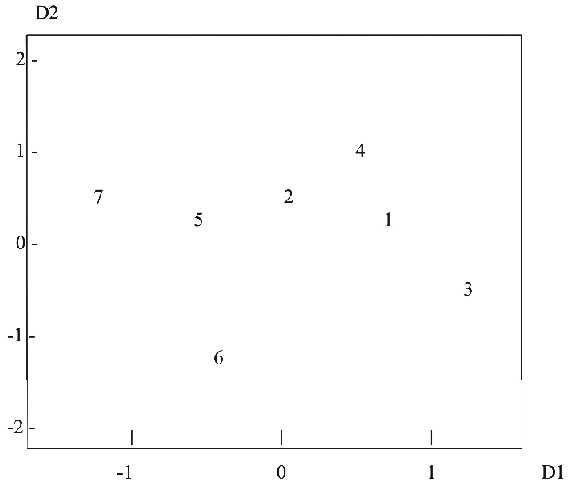
Рис. 1. Взаимное положение субпопуляций зубров, полученное методом многомерного шкалирования на основе генетических расстояний по 50 иммунологическим и 4 биохимическим полиморфным генетическим маркерам:
(1-3) кавказско-беловежские зубры из (1) ПТЗ, (2) ОГЗ и (3) Ивано-Франковска;
(4-5) беловежские зубры из (4) ПТЗ и (5) Беловежской пущи;
(6-7) горные зубры из (6) Кавказского заповедника и (7) Нальчикского охотхозяйства.
Для сравнения современных и вымерших популяций зубров (для которых невозможен анализ генетических маркеров) мы использовали фены черепа, которые могут служить аналогами генов и исследоваться такими же методами. Положение популяций зубров в пространстве двух первых измерений, полученное методом многомерного шкалирования представлено на Рис. 2. Можно видеть, что три современные популяции зубров близки друг к другу и к зубрам Беловежской пущи прошлого и начала нынешнего века, причем современные беловежские зубры несколько ближе к последним, чем кавказско-беловежские. Вместе с тем, как и при анализе генетической дифференциации среди современных субпопуляций, можно видеть относительно небольшие различия всех современных популяций по сравнению с дифференциацией вымерших популяций как кавказского, так и беловежского подвидов.
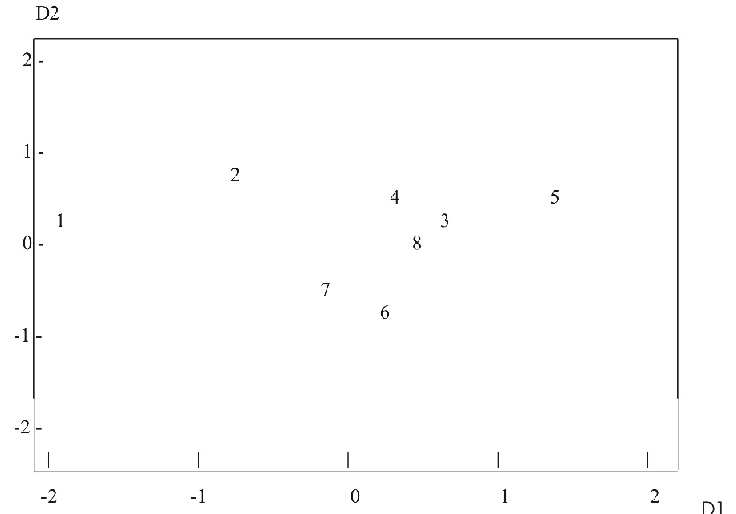
Рис. 2. Взаимное положение субпопуляций зубров, полученное методом многомерного шкалирования на основе генетических расстояний по фенам черепа:
(1) B.b.caucasicus XVIII--начала XIX века (из Северной Осетии), (2) B.b.caucasicus конца XIX--начала XX века из Кубанской царской охоты;
(3-5) B.b.bonasus: (3) из Беловежской пущи второй половины XIX века, (4) из Беловежской пущи начала XX века, (5) из Гатчинского зверинца конца XIX--начала XX века;
(6) кавказско-беловежские зубры из ПТЗ, (7) кавказско-беловежские зубры из ОГЗ и (8) беловежские зубры из ПТЗ.
Анализ признаков, характеризующих флуктуирующую асимметрию
Рядом исследований, проводившихся на разных видах животных, показано, что уровень флуктуирующей асимметрии является чувствительным интегральным показателем состояния популяции (см. Захаров, 1987). В этой связи мы провели сравнительный анализ асимметрии ряда морфологических признаков черепа в выборках из современных и исчезнувших популяций зубров.
Для оценки состояния популяций уровень флуктуирующей асимметрии был оценен в следующих выборках зубров:
1. Кавказские зубры (B. b. caucasicus), жившие на Кавказе в 1886-1912 гг.
2. Зубры Беловежской пущи (B. b. bonasus), жившие в первую половину XIX века
3. Зубры Беловежской пущи (B.b. bonasus), жившие в конце XIX-начале XX веков
4. Беловежские зубры (B.b. bonasus), из Гатчинского питомника (1861-1917 годы)
5. Современные зубры кавказско-беловежской линии, из коллекции Окского заповедника, собранной Е.Г.Киселевой в 1956-1996 гг..
6. Современные зубры беловежской линии, жившие в Центральном Зубропитомнике Приокско-Террасного заповедника в 1946-1995 гг.
7. Современные зубры кавказско-беловежской линии, жившие в ЦЗП Приокско-Террасного заповедника в 1946-1995 гг.
8. Современные зубры Кавказского заповедника горной линии предложенной в качестве подвида (B. b. montanus) за 1968-1994 годы.
Для каждой особи флуктуирующая асимметрия характеризовалась по количеству отверстий на 25 участках черепа и нижней челюсти правой и левой сторон, а также по 8 качественным признакам, характеризующим положение костей, швов и отверстий относительно друг друга. Этот набор признаков предложен А.С.Барановым (Baranov et al., 1997) и был любезно предложен нам, что делает наши исследования сопоставимыми. В таблице 13 представлены данные о среднем (в расчете на одну особь) количестве фенетических признаков с билатерально асимметричным проявлением. Три выборки из популяций современных зубров характеризуются значительным повышением уровня асимметрии по сравнению со всеми более ранними популяциями. Интересно, что наиболее симметричны, оказались зубры B.b.caucasicus, которые достоверно отличается от всех остальных, в том числе и популяций B.b.bonasus, исчезнувших одновременно с ними. Последнее можно объяснить тем, что существенный антропогенный прессинг на популяции беловежского подвида начался значительно раньше и оказал большее дестабилизирующее воздействие, сказывавшееся еще в условиях, когда виду не грозило полное исчезновение (так существуют указания на существенное сокращение численности зубра в Беловежской пуще еще в XVI веке). Кавказский подвид, вероятно, попал в условия столь жесткого воздействия позже и практически до самого момента своего вымирания сохранял высокую устойчивость онтогенеза, что выразилось в относительно более низком уровне асимметрии.
Таблица 13. Среднее на особь количество признаков с билатерально асимметричным проявлением у разных форм зубров
| Форма зубра | Объем выборки | Среднee кол-во асимметричных признаков | Достоверность различий 3 4 5 6 7 8 |
| 1. B. b. caucasicus | 31 | 4.93±0.31 | + + + + + + |
| 2. B. b. bonasus | 103 | 5.95±0.20 | + - + + + |
| 3. B. b. bonasus | 30 | 7.22±0.46 | - - + + |
| 4. B. b. bonasus | 16 | 6.49±0.58 | + + + |
| 5. B. bonasus (ОГЗ, КБ) | 75 | 68.00±0.32 | + + |
| 6. B. bonasus (ПТЗ, КБ) | 30 | 11.43±0.51 | - |
| 7. B. bonasus (ПТЗ, Б) | 33 | 11.42±0.49 | |
| 8. B. bonasus montanus | 40 | 7.48±0.47 |
(+) Различия достоверны по 5% порогу значимости;
(-) Различия недостоверны по 5% порогу значимости.
Достаточно неожиданными представляются результаты сравнения выборок из трех современных популяций: кавказско-беловежских зубров Приокско-Террасного заповедника (ПТЗ) и Окского Госзаповедника (ОГЗ), а также беловежских зубров из ПТЗ. Имея в виду, что беловежская линия происходит от меньшего числа основателей и характеризуется более высоким уровнем инбридинга, обеднения генетического полиморфизма и даже некоторыми признаками вырождения (Сипко и др., 1989; 1991; 1993; 1994; 1995; 1996), мы вправе были бы оживать у последней большего уровня асимметрии (именно такой результат был получен при сравнении зубров беловежской линии из Польши с зубрами ОГЗ (Baranov et al., 1997). Наши данные подтверждают, что по этому параметру кавказско-беловежские зубры из ОГЗ значимо стабильнее, чем беловежские зубры из ПТЗ при крайне незначительных отличиях от кавказско-беловежских зубров из ПТЗ. В настоящее время мы не можем найти объяснение малым различиям двух выборок из ПТЗ. Наиболее вероятной причиной можно считать межлинейное скрещивание и тот факт, что практика раздельного разведения линий в ПТЗ привела к тому, что эффективная численность на протяжении многих поколений для каждой из линий была ниже по сравнению с популяцией Окского заповедника. Однако, для решения этого вопроса нужны дополнительные данные; здесь мы можем лишь обратить внимание на важность дальнейших исследований.
Сравнение наших результатов с полученными ранее Барановым (Baranov et all, 1997) показывает их качественное совпадение. Как и в нашем исследовании, выборка из ОГЗ показала статистически значимые отличия от зубров беловежской линии, однако и в том, и в другом случае среднее количество асимметричных признаков на особь у нас несколько выше. Во всяком случае, несомненно, совпадение наших выводов относительно существования отличий кавказско-беловежских зубров из ОГЗ от зубров беловежской линии.
Перспективы создания вольных популяций зубра
Как уже обсуждалось, только популяция с эффективной численностью (Ne) в 500 особей гарантирует существование вида длительное время. Для реализации этого правила требуется большее, чем поддержание группы животных подобной численности, так как фактическое число взрослых, половозрелых и участвующих в размножении особей может отличаться от общей численности популяции (N) полученной в результате учета. У видов с гаремным характером размножения, к которым и относится зубр, эффект относительно малого числа размножающихся самцов заключается в том, что существенно увеличивается минимальное фактическое число особей, требуемое для достижения желаемой эффективной численности. Для таких животных обычна ситуация, когда отношение N/Ne составляет 3-4 (Soule, 1980). Это соответствует общей численности группировки в 1500-2000 голов, которая только и может гарантировать сохранение вида в долговременном плане. И ставя перед собой задачу спасения и сохранения зубра необходимо организовывать разведение этого вида так, чтобы все совместные усилия специалистов были направлены на создание популяций подобного размера.
Общее число зубров в мире не достигает трех тысяч, а высокая смертность и низкие темпы прироста поголовья не позволят в краткие сроки добиться создания подобной популяции. По этой причине для достижения подобной цели требуется скоординированное действие большинства центров разведения зубров Европы, и в течение многих лет.
Обсуждая вопрос о числе популяций, необходимых для сохранения этого вида можно говорить о том, что во избежание последствий эпидемий или других форс-мажорных обстоятельств нужно иметь не менее двух и значительно изолированных популяций, обеспечивающих гарантированное поддержание генофонда. Так как современное число зубров невелико, а темпы прироста популяций составляют примерно 3-6% в год и имеют тенденцию к снижению рассчитывать, что в ближайшие десятилетия эта задача будет решена без концентрации всех племенных ресурсов зубра мира на создание не более двух популяций, более чем легкомысленно.
Районы, где расположены Скнятинское, Великоозерское охотничьи хозяйства и Фоминский заказник и т.п., имеются небольшие группы зубров, и не могут обеспечить долговременное существование крупных популяций, нами не рассматриваются. Также неэффективно было бы распылять силы, средства и племенное поголовье для создания популяции на территории, которая может вместить не более 100-200 особей.
Известно, что в зависимости от условий обитания на одну голову требуется от 2 до 10 км2. Используя средние данные по допустимой плотности нужно говорить о территории в 9-12 тысяч км2. Практически вся территория Русской равнины сильно трансформирована многовековой деятельностью человека (Попадюк и др., 1994). Реально, в наше время, лесные массивы сохраняются только на территориях с наиболее низкой продуктивностью, то есть на бедных почвах или в местах с избыточным увлажнением. При этом большинство лесных массивов представляют собой культурные или окультуренные насаждения, от которых предполагается получение продукции. Можно даже считать большую часть этих лесов сельскохозяйственными угодьями со своим режимом использования, где воздействие копытных нежелательно. Необходимо подчеркнуть следующую особенность биологии зубра имеющую сходство среди нашей фауны, пожалуй, только с овцебыком. Этот вид живёт в основном, осёдло образуя постоянные группы числом в 12-30 особей. И нетрудно представить размеры ущерба с\х культурам и лесным посадкам, когда их посетит группа зубров общим живым весом до 12 тонн! А значит, территория, предназначенная для расселения зубра, не должна рассматриваться как объект промышленного лесоразведения, и должна находится по возможности изолированно от сельскохозяйственных площадей (это обычно охраняемые природные территории). Общие требования к району создания вольной популяции зубра будут следующие:
1. Соответствовать биологическим потребностям этого вида.
2. Представлять собой компактно расположенный природный комплекс, пригодный для обитания от 1000 и более особей.
3. Иметь надежную охрану для зубра.
4.Иметь законодательные гарантии о неизменности охраняемого статуса этой территории на многие десятилетия.
5.Найти территории отвечающие подобным требованиям достаточно сложно. Их нет на территории западной Европы, а в восточной Европе такими возможностями располагает только Россия.
В первую очередь это национальный парк "Орловское полесье" который имеет 360 км2 лесопокрытой территории, протянувшейся более чем на 50 км вдоль границы с Калужской областью. Он является окраиной лесного района площадью более 1000 км2. На северо-западе к парку примыкает заповедник "Калужские засеки" и еще один крупный лесной массив. Южнее расположен заповедник "Брянский лес", который является частью единого и компактного лесного массива расположенного по левому берегу р. Десны с примерными размерами 100х30 км. Расстояние между местами обитания зубров составляет: Брянский лес - Конотопское стадо - 150 км Брянский лес - Даневское стадо - 260 км Брянский лес - Орловское полесье - 150 км. Охрана территорий заповедников, национального парка и их охранных зон хорошо налажена. Но лесной практически единый массив, разделенный только административными границами, не является препятствием для движения зубров и браконьеров, а со стороны Калужской области вообще нет никаких служб охраны, позволяющих гарантировать охрану зубра. Программу по созданию популяции зубра в "Орловском полесье" можно выполнять только вместе с проведением необходимых мероприятий по организации охраны зубра на смежной территории Калужской области.
По всему комплексу условий этот район можно рассматривать как оптимальный для создания крупной саморегулирующейся популяции зубра. А национальный парк имеет все условия, чтобы стать ядром формирования крупной и саморегулирующийся популяции зубров. Но только в том случае, когда работа по созданию популяции будет продолжена в течение нескольких десятков лет и получит дополнительное финансирование.
По непроверенным (опросным) данным, имеются сведения о встречах с зубрами, зашедшими с сопредельной территории Украины в районы, примыкающие с юга к границам заповедника «Брянский лес». Стали появляться зубры у заповедника «Калужские засеки». Завоз зубров в заповедник «Брянский лес» окончился неудачей, чему основная причина - плохая организация этой работы. На территорию национального парка «Орловское полесье» в 1996 - 2002 годы было завезено 67 особи зубра. Уже два года отмечались отёлы и сейчас в полесье 78 зубров. Наибольший по численности отел был в 2002 году и составил 8 телят (по сообщению В.Д. Казмина). Принимая во внимание то, что в числе этой популяции уже более 40 взрослых самок можно также говорить о низком репродуктивном потенциале этого вида.
По нашим расчетам дополнительные завозы зубров в этот регион, целесообразно продолжать до достижения популяцией общей численности в 400 особей. Так как только с этого времени уже будет маловероятно или вообще невозможно завести группу зубров сравнимую по числу с годовым приростом, а значит и существенно повлиять на популяцию. А достижение численности популяции в 400 особей можно ожидать с учетом ежегодного завоза 5-10 особей к 2020 году в лучшем случае. В этом случае на основе проведенного нами математического моделирования эта популяция достигнет рекомендуемой численности в 1,5 тысячи особей к 2033-2037 годам.
Второе место, которое наиболее целесообразно использовать для создания полноценной популяции это Усть-Кубенское охотничье хозяйство Вологодской области. Вологодская область находится на значительном расстоянии от Орловской области (около 800 км), что исключает возможность одновременного наступления в обоих районах эпидемий, стихийных бедствий и др. форс-мажорных обстоятельств. Пригодность этого района для обитания зубра подтверждается тем, что без зимней подкормки с 1991 года здесь успешно адаптировались и регулярно размножаются зубры. Эти места в историческом прошлом занимали широколиственные леса (Попадюк и др., 1994), имеются данные об обитании зубров в этом регионе (Кулагин, 1919) и соседних, включая Финляндию (Соколов, 1959). Территория хозяйства и окрестности имеют обширные площади бывших сельскохозяйственных угодий и лесные вырубки. Богатые кормовые ресурсы и большая площадь около 120х100 км с чрезвычайно редким населением позволяют рассчитывать на обитание здесь более 1000 особей. Отсутствие развитой дорожной сети и надежная охрана практически гарантируют безопасность зубра. От выпущенных 5 особей (из них две самки в 1994г) получено 4 теленка, причем из них 3 самца, что вызвано, скорее всего, генетическими причинами. Репродуктивный успех этого стада стал бы выше, если вместо 3 самок беловежской линии использовали другую линию. Так как адаптационные возможности животных беловежской линии не вполне соответствует условиям этого региона.
Для освежения крови и нормализации демографической ситуации срочно необходим завоз как минимум 2-5 самок кавказско-беловежской линии, подобранных по родословным, генетическим маркерам и возрасту. Для того чтобы можно рассчитывать на создание саморегулирующейся демографически и генетически устойчивой популяции, где минимизированы дальнейшие потери генетического разнообразия, необходимы также как и в «Орловском полесье» завозы животных из Российских и зарубежных пунктов их содержания на постоянной основе.
Благоприятные для обитания зубра, районы северного Кавказа, в настоящее время являются зоной политической и экономической нестабильности. И привозить туда в ближайшее десятилетие столь редких животных было бы неразумно. С другой стороны в Кавказском государственном заповеднике с 1940 года существует самая крупная и старейшая вольно живущая популяция зубра. Эта форма получила в настоящее время статус подвида и имеет наибольшее генетическое и генеалогическое разнообразие по сравнению с другими вольными популяциями. Её численность превышала 1500 особей, что гарантировало формирование естественной демографической и генетической структуры популяции. В настоящее время число горных зубров составляет 450 особей, хотя в районе заповедника и прилегающих охраняемых территориях могут обитать до 5 тысяч особей (Yasan et al., 1987). И проблема состоит только в организации действенной охраны и рационального использования.
В качестве страховочного варианта, на перспективу, следует рассматривать Сибирь или Дальний восток. Это районы, где достаточное количество территорий отвечающих указанным выше параметрам. В пользу рассмотрения этих регионов говорит и тот факт, что еще в VII веке нашей эры в южной Сибири (Верещагин, Барышников, 1985) обитали зубры. На полярном Урале, завезенные в качестве эксперимента бизоны, несколько лет жили без подкормки. Также много лет функционирует питомник зубров на Алтае.
Нельзя обойти вниманием тот факт, что охрана ради охраны не может продолжаться длительное время и только организация хозяйственного использования зубра (Павлов, 1999), к примеру трофейные охоты позволит иметь перспективу в расселении этого вида.
Выводы
1. Беловежская линия существенно уступает кавказско-беловежской по всем рассмотренным витальным, генетическим и морфологическим характеристикам;
2. На примере зубропитомника Окского и Приокско-Террасного заповедника за последние 30 лет прослеживается негативная динамика параметров плодовитости и жизнеспособности; эти выводы можно экстраполировать и на зубров из других питомников.
3. Генофонд зубров в России значительно обеднен и в имеющихся питомниках происходит потеря генетического разнообразия;
4. В генотипах современных зубров очень неравномерно и не полно представлен генетический пул родоначальников;
5. Генетическая дифференциация современных зубров очень низка, ниже, чем у вымерших популяций конца прошлого-начала нынешнего века; она сопоставима с дифференциацией стад одной породы крупного рогатого скота (а не разных пород); беловежская линия генетически близка кавказско-беловежской линии зубров и может рассматриваться, как часть последней;
Список цитируемой литературы
Абилов А.И., Эрнст Л.К., Срекозов Н.И., Кононов Б.П., Сипко Т.П. Методические рекомендации по получению гибридов путем осеменения домашних коров (Bos taurus) эпидидимальным семенем диких зубров (Bison bonasus). ВИЖ, Дубровцы 1994, 26 с.
Башкиров И.Г. Кавказский зубр. М: Гл. Упр. по заповедникам, зоопаркам и зоосадам. 1939, С. 3-72.
Белоусова И.П. Влияние инбридинга на жизнеспособность зубров в питомниках России (Bison bonasus) // К вопросу о возможности сохранения зубра в России. Сб. трудов. Пущино. 1993. С. 29-43.
Буневич А.Н. Воспроизводительные показатели популяции зубров Беловежской пущи //Редкие виды млекопитающих России и сопредельных территорий. Москва, 1997, с. 17.
Буневич А.Н. Беловежская пуща. Исследования. Анализ формирования популяции зубра в Белорусской части Беловежской пущи. Вып. 11. Брест: Изд-во С. Лаврова, 2003 С. 178-205.
Верещагин Н.К., Барышников Г.Ф. Вымирание млекопитающих в четвертичном периоде северной Евразии. \\ Млекопитающие северной Евразии в четвертичном периоде. Труды зоологического института АН СССР, Ленинград, 1985, Том 131, С. 3-38.
Животовский Л.А. Популяционная биометрия. М.: Наука, 1991, 272 с.
Заблоцкий М.А. Тридцать пять лет разведения зубров и из гибридов в зоопарке Аскания-Нова. В кн. Кавказский зубр. Москва, 1939, с 73-137.
Захаров В.М. Асимметрия животных. М.: Наука, 1986, 215 с.
Карцов Г.П. Беловежская пуща. СПб.: Изд-во. Маркса. 1903, 419 с.
Калугин С.Г. Зубры в естественных условиях Кавказского заповедника. Труды Кавказского государственного заповедника вып. IV.. Майкоп 1958, С.4-37.
Калугин С.Г. Разведение зубров в Кавказском заповеднике. Труды Кавказского государственного заповедника вып. VIII.. Краснодар 1965, С.129-155.
Кириков С.В. Распространение зубра на территории Советского союза в XI-XX вв. \\ Зубр. Морфология, систематика, эволюция, экология. М., Наука, 1979, С.476-486.
Корочкина Л.Н. Показатели размножения зубров Беловежской пущи // Беловежская пуща, Минск: Урожай, 1971, вып. 4 с. 176-184.
Кулагин Н.М. Зубры Беловежской пущи изд. МНИ., М., 1919, 166 с.
Кренке А.Н.,Чернавская М.М., Браздил Р. и др. Изменчивость климата Европы в историческом прошлом. М., Наука, 1995, 224 с.
Павлов М.П. Акклиматизация охотничье-промысловых зверей и птиц в СССР. Часть III:Копытные. Киров.1999, 666 с.
Попадюк Р.В., Чистякова А.А., Чумаченко и др.; Под ред. Смирновой О.В. Восточноевропейские широколиственные леса. М.: Наука, 1994, 354 с.
Раутиан Г.С., Сипко Т.П., Уханов С.В., Генетический полиморфизм зубра. Материалы 1-го съезда Вавиловского общества генетиков и селекционеров (ВОГИС).(Саратов, 20-25 декабря 1994г.) //Генетика, т.30, 1994, с. 131.
Раутиан Г.С., Калабушкин Б.А., Немцев А.С. Новый подвид зубра В. b. Montanus ssp. Доклады академии наук. Общая биология. 2000, т. 375, № 4, С. 563-567.
Рекомендации, выработанные на совещании экспертов ФАО/ЮНЕП по восстановлению лошади Пржевальского в Монголии // Сборник мат. совещ. экспертов ФАО/ЮНЕП СССР 29-31 мая 1985 г. М.: Центр Междунар. Проект. ГКНТ. 1988, С. 8-26.
Розанов М.П. Зоологическая экспедиция Главнауки в Кавказский заповедник. Охрана природы, 1928, № 3.
Сатунин К.А. Кавказский зубр. Естеств. и геогр., февраль, 1898, C. 6-8 .
Северцов С.А. Видовые константы размножения Беловежского зубра и динамика населения этого вида. Тр. Института эволюционной морфологии. Т. III, вып.1, изд. АН СССР, М., 1940, С.3-32.
Соколов И.И. Фауна СССР Млекопитающие, т. 1., вып. 3, М.-Л., Изд. АН СССР, 1959, 640 с.
Силантьев А.А. Зубр (Bison bonasuis L.). Естеств. производит. сил. России, т. VI, отд. 1. Млекопитающие; птицы. Изд. Академия наук, Петр.,1919, с. 132-145.
Сипко Т.П., Черкащенко В.И. Генетический анализ зубров в СССР // Современные проблемы Красной книги СССР. М. 1989. С. 31-37.
Сипко Т.П. Об определении статуса горных зубров Северо-Западного Кавказа./ Материалы научно-практической конференции, посвященной 50-летию регулярных исследований в Беловежской пуще (19-21 декабря 1989г.). Минск, Каменюки. 1990, с. 154-156.
Сипко Т.П., Абилова А.И., Шурхал А.В., Ротт Н.Н. Создание криобанков спермы зубра как метод консервации генетического полиморфизма вида // Генетика, 1993, Т. 29, N 3, с. 2051--2056.
Сипко Т.П., Белоусова И.П. Анализ состояния генофонда зубра в питомниках России.//К вопросу о возможности сохранения зубра в России. Пущино, 1993. с. 56-62.
Сипко Т.П., Удина И.Г., Бадагуева Ю.Н., Сулимова Г.Е. Сравнительная характеристика полиморфизма ДНК гена каппа-казеина у представителей семейства Bovidae. / Генетика, 1994, Т. 30, N 2, c.225-229.
Сипко Т.П., Раутиан Г.С., Удина И.Г., Уханов С.В., Берендяева З.И. Изучение полиморфизма групп крови у зубров (Bison bonasus) // Генетика, 1995, Т. 31, N 1, с. 93-100.
Сипко Т.П., Г.С.Раутиан, И.Г.Удина, Т.А. Ракицкая. Полиморфизм биохимических маркеров зубра (Bison bonasus). // Генетика, 1996, Т. 32, N 3, с. 400-405.
Сипко Т.П., Раутиан Г.С. Динамика факторов роста численности зубров // Редкие виды млекопитающих России и сопредельных территорий. М., 1997 с. 88.
Сипко Т.П., Ломов А.А., Банникова А.А., Медников Б.М. Оценка степени генетической дивергенции некоторых полорогих методом рестриктазного анализа ДНК .// Цитология и генетика. 1997. Т. 31. N 4, c. 76-81.
Сулей Э.С. Пороги для выживания: поддержание приспособленности и эволюционного потенциала. Conservation biology. An Evolutionary – Ecological Perspective. Edited by Soule M.E. and Wilcox B.A, 1980 by Sinauer Associates, Inc. Publishers Sunderland, Massachusetts. P. 176-197.
Удина И.Г., Соколова С.С., Сипко Т.П., Сулимова Г.Е. Сравнительная характеристика полиморфизма ДНК локусов DQB и DRB главного компонента гистосовместимости у представителей семейства Bovidae // Генетика. 1994. Т.30. N 3. С. 356-360.
Усов С.А. Зубр. Записки Русского общ. Акклиматизации; изд. Москв. Зоологического сада, М., 1865 (Перепеч. В Соч. С.А. Усова, т.1; М.,1888).
Флеров К.К. Ситематика и эволюция \\ Зубр. Морфология,систематика, эволюция, экология. М.,Наука, 1979, С. 9-127.
Фортунатов Б.К. В поисках зубра. (Заметка о фауне Кавказского Гос. Заповедника, III). Природа и соц. Хоз.-о, т. V, 1932. C. 12-15.
Франклин Ян Р. Эволюционные изменения в небольших популяциях. Conservation biology. An Evolutionary – Ecological Perspective. Edited by Soule M.E. and Wilcox B.A, 1980 by Sinauer Associates, Inc. Publishers Sunderland, Massachusetts. P. 160-176.
Юргенсон П.Б., Заблоцкий М.А. Государственная племенная книга зубров и бизонов. Т.1, изд. МСХ СССР, Москва 1956, с. 1-114.
Шапошников Ч.Г. Кавказский гос. Заповедник. Охрана природы. 1928, №1,2,6.
Шильдер В. Кубанская охота великого князя Сергея Михайловича в 1894 г. Природа и охота, 1895, № 5,7,8.
Baranov A.S., Pucek Z., Kiseleva E.G., Zakharov V.M. Developmental stability of skull morphology in European bison, Bison bonasus // Acta Theriol. 1997, Suppl. 4. (Developmental homeostasis in natural populations of mammals: phenetic approach. Zakharov V.M., Yablokov A.V., Eds.). P. 79-85.
Baranov A.S., Zakharov V.M. Developmental stability in hybrids of European bison Bison bonasus and domestic cattle // Acta Theriol. 1997. Suppl. 4. (Developmental homeostasis in natural populations of mammals: phenetic approach. Zakharov V.M., Yablokov A.V., Eds.). P. 87-90.
European Bison Pedigree Boor 1953-2000 (EBPB) Editor Raczynski J. Bialowieza, 2000, 65 p.
Ferguson M.M., Drahuschak L.R. Disease risistance and enzyme heterozygosity in raindow trout // Heredity. 1990. N 64. P.413-417.
Frankel O.H. & M.E.Soule,. Conservation and Evolution. Cambridge University Press, Cambridge, England. Frankel J.R., 1980. Evolutionary change in small populations, p. 135-149. In M.Soule and B.Wilcox (eds.), Conservation Biology: An Evolutionary-Ecological Perspective. Sinauer Associates, Sunderland, Massachusetts.
Hartl G.B., Pucek Z. Genetic depletion in the European bison (Bison bonasus) and the significance of electrophoretic heterozygosity for conservation // Conservation biology, 1994, v. 8, N 1, p. 167-174.
Krasinski Z.A., Bunevich A.N., Krasinska M. Charakterystyka populacji zubra nizinnego w polskiej i bialoruskiej czesci Puszczy Bialowieskiej/ Parki Narodowe i Rezerwaty Przyrody. Tom 13, N 4, 1994, p.25-67.
Nevo E., Noy R., Lavi B., et al Levels of genetic diversity and resistance to pollution in marine organisms //FAO Fish. Rep. 1985. N 352, suppl. P. 175-182.
O`Brien S.F., Evermann J.F. Interactive influence of infectius disease and genetic diversity in natural populations // Trends Ecol. Evol. 1988. N 3. P.254-259.
Olech W. Analysis of inbreeding in European bison // Acta theriol., 1987. V.30, P.373-387.
Olech, W., The Participation of Ancestral Genes in the Existing Population of European Bison // Acta Theriol., 1989, v. 34, p. 397-407.
Parsons P.A. Environmental stresses and conservations // Ann. Rev. Ecol. Syst. 1989. Vol. 20. P. 29-49.
Pucek Z. History of the European bison and problems of its protection and management. In: Global trends in wildlife management. Krakow-Warszawa 1991, p. 19-39.
Pucek Z., Seal U.S., Miller P. Population and habitat viability assessment for the European bison //A Collaborative Workshop in Wolinski National Park Miedzyzdroje, Poland 26-29 June 1995. 193 p.
Sennner J.W., 1980. Inbreeding depression and the survival of zoo populations, pp. 209-244. In: M.E.Soule and B.A.Wilcox (eds.), Conservation Biology: An Evolutionary-Ecological Perspective. Sinauer Associates, Sunderland, Massachusetts.
Sipko T.P. at el. Conservation of Genetic Material from Endangered and Economically Important Ungulate Species in the Establishment of Cryobanks //Physiology and Gen. Biol. Rev., Harwood Acad. Publ. 1997. v. 13., part 3, p. 1-99.
Soule M.E., Wilcox B.A. (Edited) Conservation biology.Sinauer Associates, Inc. Publishers Sundderland, Massachusetts, 1980. 430 p.
Yasan Y.P., Sipko T.P, Kashtanov S.N., Nemtzev A.S. Analysis of Bison bonasus populations according to markers.// Abst. XYIII th. Congress of International Union of Game Biologists.- Krakow, Poland, 1987. p. 217-218.
Westberg G. Ueber die Verbreitung des Wisent im Osten des europaisch-asiatischen Kontinents. Arbeit. Des Naturf.-Vereins zu Riga, Neue Folge, H. 9; 1899. P. 17-41.
Westberg G. Einiges uber Bisone und die Verbreitung des Wisents im Kaukasus. Festscnrift des Naturf.-Vereins zu Riga in anlass seines 50 jahrigen Bestehens. Riga, 1895. 23 p.
Wroblewski K. Zubr Puszczy Bialowieskiej. Wyd. Polskie. Poznan: 1927., 232 p.